![]()
![]()
Los patrones resultantes de la evolución son analizados básicamente por dos disciplinas: la sistemática y la biogeografía.
Es la disciplina que estudia la clasificación de los seres vivos. También se la conoce como taxonomía, aunque algunos autores prefieren reservar este último término para cuestiones teóricas, referidas a los principios y procedimientos clasificatorios, mientras que emplean el nombre de sistemática para un campo del conocimiento más amplio.
El objetivo principal de la investigación sistemática es obtener una clasificación de los seres vivos estudiados. ¿Para qué se construyen clasificaciones? Porque permiten manejar una gran cantidad de información y recuperarla fácilmente. Cuando se dice que una determinada especie pertenece a la clase Mammalia —como se denomina el grupo de los mamíferos— automáticamente se asume que tendrá glándulas mamarias, pelos y algunas características óseas particulares, sin necesidad de detallarlas. Además, las clasificaciones permiten predecir información desconocida. Por ejemplo, si un conjunto de especies comparte cinco caracteres y se descubre una nueva especie que se asignaría a ese grupo, aunque sólo puedan analizarse cuatro de estos caracteres, es posible predecir que esa especie también tendrá el quinto carácter no observado.
En la historia de la sistemática se han desarrollado varios métodos clasificatorios. Los principales son la taxonomía gradista, la fenética y la cladística.
La taxonomía gradista o evolucionista se basa en la idea de agrupar taxones considerando, además de las relaciones genealógicas, el grado de diferencia entre los mismos. Los taxónomos gradistas admiten grupos parafiléticos o grados en sus clasificaciones, si hay un intervalo morfológico notorio entre un grupo monofilético y la serie de taxones que lo anteceden, si el grupo monofilético ocupa una zona adaptativa diferente que la serie de taxones que lo anteceden, y si hay una mayor riqueza de especies en el grupo monofilético, en relación con la relativa pobreza de la serie de taxones que lo anteceden. Uno de los casos más conocidos es el de los reptiles. Aunque este taxón no incluye todos los descendientes de su ancestro común, los gradistas consideran que el grado de diferencia que poseen con las aves y los mamíferos justifica que se los mantenga como tres taxones separados. La metodología gradista es muy heterogénea. En 1961, George G. Simpson (1902-1994) resumió la práctica taxonómica gradista, aunque no detalló procedimientos analíticos precisos.
La taxonomía fenética o numérica se basa en la idea de agrupar taxones por su similitud global. Luego de analizar un gran número de caracteres y evaluar su distribución en los taxones analizados, se obtienen coeficientes que expresan las relaciones entre estos últimos y se construye un fenograma. En este diagrama los taxones se agrupan de acuerdo con su parecido global. Un problema con la fenética es que las estimaciones basadas en similitud global pueden "mezclar" caracteres que se aplican a diferentes niveles de universalidad del problema examinado. Otro problema es que es posible obtener diferentes agrupamientos a partir de la misma matriz de datos, si se emplean distintos algoritmos, por lo que no se logra la objetividad y estabilidad buscadas. Pese a sus deficiencias, la taxonomía fenética constituyó una evidente mejora en relación con la taxonomía gradista, de algún modo tendiendo a disminuir las decisiones subjetivas. Actualmente la taxonomía fenética ha sido abandonada casi por completo, salvo por su uso para resolver algunos problemas por debajo del nivel de la especie.
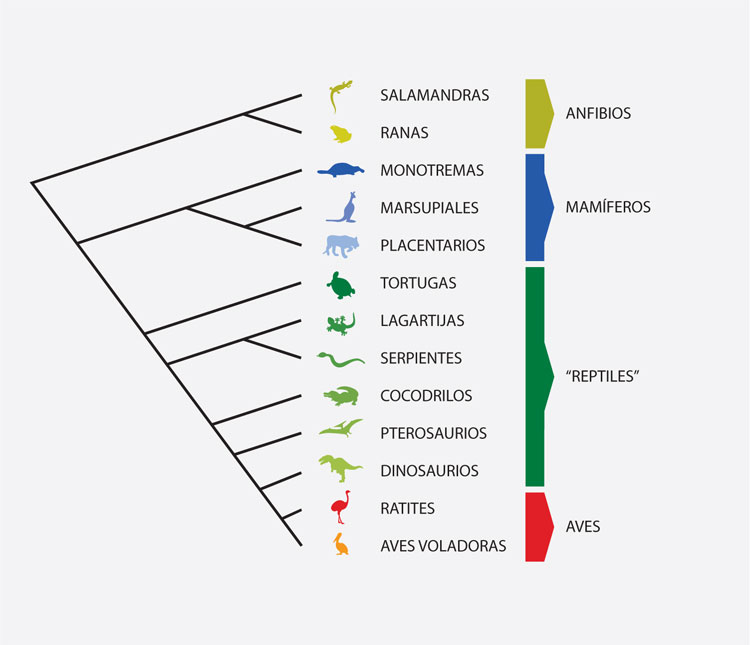
La sistemática filogenética o cladística considera que la clasificación de los seres vivos debe basarse en el reconocimiento de sus relaciones genealógicas, a través de un cladograma (como el que se presenta arriba). En éste se detallan los caracteres que justifican cada uno de los agrupamientos taxonómicos, denominados clados o grupos monofiléticos. Los clados son los únicos grupos que reconocen como válidos los taxónomos cladistas. Así, por ejemplo, tiene sentido reconocer a la clase Insecta como un grupo monofilético natural, pero no puede reconocerse un grupo que reúna a los insectos carentes de alas, pues las alas se han perdido varias veces independientemente en varios clados del grupo. El fin fundamental de un análisis cladístico es obtener un cladograma, para lo cual deben seguirse estos pasos: 1] seleccionar los taxones que serán las unidades del estudio; 2] seleccionar los caracteres que brindarán la evidencia sobre las relaciones genealógicas de los taxones estudiados; 3] descubrir las relaciones genealógicas de los taxones analizados y volcarlas en un cladograma, y 4] traducir las relaciones genealógicas del cladograma en una clasificación formal.
Un taxón es un grupo de seres vivos considerado como una unidad de cualquier categoría en una clasificación. Por ejemplo, Homo sapiens, Primates o Animalia. No debe confundirse un taxón con la categoría que se le asigna; es decir, el rango o nivel en una clasificación jerárquica, pues a un mismo taxón se le pueden asignar categorías diferentes, de acuerdo con el autor que lo analice. Por ejemplo, Homo sapiens es una especie, Primates es un orden y Animalia es un reino.
Un carácter es cualquier atributo observable en un taxón, que permite distinguirlo de otro. Cabe aclarar que no toda la variación que se encuentra en los seres vivos posee valor taxonómico. Algunos ejemplos de variación carente de valor taxonómico son la variación individual en el tiempo (organismos que varían con la edad o con las estaciones del año), la variación social (en los insectos sociales, como las abejas, las hormigas y las termitas, en que hay sistemas de castas), la variación ecológica (organismos que cambian algunas de sus características en relación con el ambiente, como plantas que crecen más achaparradas en climas áridos, o parásitos que cambian su aspecto según el huésped), la variación traumática (variaciones en la morfología debidas a accidentes o inducidas por la acción de parásitos) y la variación asociada con el sexo (cuando existen diferencias entre los organismos masculinos y femeninos de una misma especie).
En relación con los tipos de caracteres taxonómicos, existen diferencias obvias entre distintos tipos de organismos. No serán los mismos caracteres los que emplee un botánico o un zoólogo, e incluso los que este último emplee también variarán de acuerdo con el taxón estudiado, pues no es lo mismo estudiar mamíferos que insectos. A pesar de esta enorme diversidad, podemos ensayar una clasificación de los tipos de caracteres: 1] morfológicos: externos, anatómicos (de la morfología interna), embriológicos, palinológicos (caracteres de los granos del polen), citológicos (caracteres de la célula) y ultraestructurales; 2] fisiológicos: constantes de crecimiento, regulación de la temperatura, entre otros; 3] químicos: presencia de pigmentos y alcaloides, ácidos nucleicos, proteínas y lípidos; 4] etológicos: pautas de comportamiento durante el cortejo, canto, etc.; 5] ecológicos: hábitat, parásitos, alimentación, etc.; 6] genéticos: esterilidad entre especies afines, y 7] geográficos: referidos al área de distribución.
Las diferentes alternativas de un carácter taxonómico se denominan estados. Por ejemplo, para el carácter "presencia de pelo" los estados pueden ser "presencia" o "ausencia", mientras para el carácter "número de manchas" los estados pueden ser "dos", "cuatro", "seis", etcétera.
Con el objeto de lograr una mayor precisión, en la sistemática filogenética se han propuesto diferentes términos para referirse a los estados de carácter:
1. Estado plesiomórfico: es aquel que surge primero en el tiempo, pues se halla o se infiere que se hallaba en el antecesor del grupo en estudio.
2. Estado apomórfico: es aquel que surge a partir del estado plesiomórfico, es decir, que resulta de una transformación dentro del grupo en estudio.
3. Simplesiomorfía: es un estado plesiomórfico presente en dos o más taxones.
4. Autapomorfía: es un estado apomórfico presente en un único taxón.
5. Sinapomorfía: es un estado apomórfico presente en dos o más taxones.
Cuando un carácter se desarrolla independientemente a partir de ancestros diferentes se habla de homoplasias. Si el estado homoplástico proviene del mismo estado ancestral, se habla de un paralelismo. ¿Por qué ocurre esto? Porque la evolución muchas veces actúa produciendo estructuras independientemente, quizá como adaptación a las mismas condiciones ambientales a que se ven sometidos taxones diferentes. Un caso notable lo constituyen los mamíferos marsupiales y los placentarios, en los que existen muchas formas "repetidas" en Australia y el Viejo Mundo, como "lobos", "topos" y "ratones", entre otros.
Las sinapomorfías de un grupo pueden perderse en algunos de sus descendientes, los que entonces poseen el estado plesiomórfico. En este caso se habla de reversiones. Éstas se dan porque a veces una estructura pierde su función adaptativa y así puede ser "perdida" en el curso de la evolución. Por ejemplo, las ballenas han perdido sus miembros posteriores, aunque su cintura pélvica aún señala que alguna vez los tuvieron.
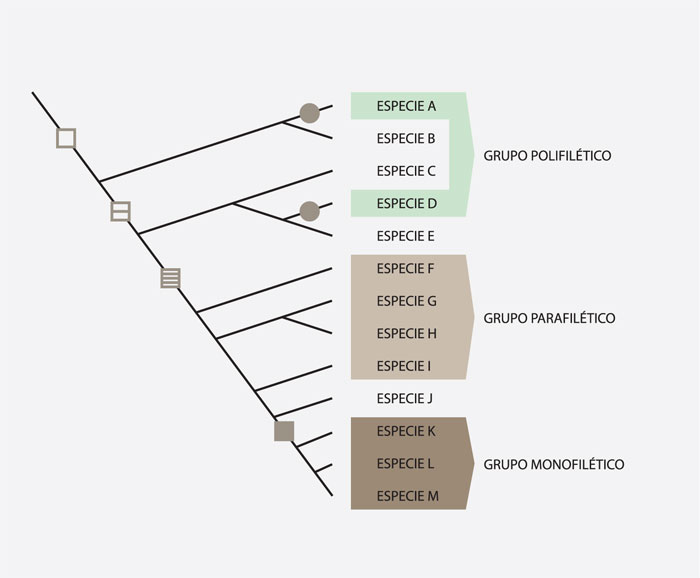
En los cladogramas es posible reconocer tres tipos de grupos diferentes: monofiléticos, parafiléticos y polifiléticos.
Los grupos monofiléticos o clados son aquellos que incluyen todos los descendientes de un ancestro común, es decir, que poseen existencia real, como resultado de la evolución, y pueden ser reconocidos por compartir una o más sinapomorfías o por poseer una combinación particular de caracteres. Los grupos parafiléticos o grados son aquellos que excluyen algunos de los descendientes del ancestro común, y están basados en simplesiomorfías. Los grupos polifiléticos son aquellos que incluyen descendientes de ancestros distintos, y están basados en paralelismos o convergencias. En un sistema filogenético, los únicos grupos que tiene cabida son los monofiléticos, pues los parafiléticos y polifiléticos son considerados artificiales.