
![]()
![]()
La evolución produce nuevas formas biológicas, es decir, da origen a la biodiversidad. La unidad básica de la gran diversidad que existe en la Tierra es la especie. Los biólogos de diversos campos de estudio generalmente utilizan el concepto biológico de especie; en este sentido, una especie es definida como una población o serie de poblaciones con individuos que pueden reproducirse y dejar descendencia fértil, es decir, son poblaciones aisladas reproductivamente de otras; esta definición se aplica a todos los organismos con reproducción sexual (la mayoría de los animales y gran parte de las plantas con flores, no puede aplicarse a organismos con reproducción asexual). Los humanos, por ejemplo, pertenecen a la especie Homo sapiens, que está integrada por muchas poblaciones dispersas alrededor del mundo. Aun cuando habitantes de distintas regiones geográficas difieren en algunos rasgos morfológicos, todos forman parte de una misma especie, todos los grupos poseen un acervo genético común y pueden reproducirse entre ellos. Pero, al mismo tiempo, existen barreras reproductivas que los separan de sus parientes más cercanos, los gorilas y los chimpancés.
La definición anterior no puede aplicarse en todos los campos, por ejemplo, las especies de fósiles sólo se describen de acuerdo con características morfológicas distintivas debido a la imposibilidad de determinar si existieron o no barreras reproductivas. El concepto de especie biológica es difícil de aplicar en organismos como las bacterias, que no poseen reproducción sexual, aunque se conocen mecanismos de intercambio de material genético, como la conjugación.
La gran diversidad de vida que existe en el planeta y la relación evolutiva entre las especies es estudiada por la sistemática. Esta disciplina estudia la diversidad como un todo, es decir, sus relaciones filogenéticas o evolutivas, ecológicas y palentológicas, organizando la diversidad en forma de clasificación jerárquica, reconociendo diversos grupos o taxones (especies, géneros, familias, clases, órdenes, phyla y reinos).
Durante el proceso evolutivo las poblaciones de una especie se van diferenciando genética, ecológica y morfológicamente. La diferenciación puede ser de tal magnitud que se produzcan barreras que impidan la reproducción. Un ejemplo claro de una barrera reproductiva se presenta en los caballos (Equus caballus) y los burros (E. asinus) que, aunque pueden cruzarse, sus descendientes (mulas o burdéganos) son estériles. La infertilidad de los híbridos puede resultar de la presencia de diferencias estructurales entre cromosomas que impidan su correcta segregación durante la meiosis o porque exista una diferencia entre genes de los progenitores.
Las especies afines se agrupan en géneros. Equus es el género al que pertenecen los caballos, burros y cebras. En tanto, el segundo nombre hace referencia a la especie: E. caballus para el caballo; E. asinus para el burro; E. burchelli, E. grevyi y E. zebra para las tres especies vivientes de cebras; y E. quagga, que corresponde a una cebra ya desaparecida. El género Equus, junto con otros géneros de caballos extintos como Hyracotherium, Merychippus o Pliohippus, se agrupan en una familia denominada Equidae. Junto con las familias Tapiridae (tapires) y Rhinocerontidae (rinocerontes), constituyen el orden Perissodactyla.
Es muy importante subrayar que las especies vivientes descienden de otras que han existido en el pasado. Al estudio de las relaciones entre especies vivientes y sus ancestros se denomina filogenia. Los grandes avances en la biología molecular permiten reconstruir la filogenia de muchos grupos de organismos. Para ello se analiza la secuencia de bases de genes nucleares y mitocondriales —o del cloroplasto en el caso de los eucariontes o del material genético de los procariontes—, y se determina qué diferencias o similitudes hay entre ellos. El principio es muy simple: si aparecen muchas divergencias en la secuencia de nucleótidos del ADN de dos especies, se deduce que tienen una relación filogenética muy lejana, y viceversa
¿Cómo se producen tales diferencias en la secuencia del ADN de un gen en distintos organismos? Básicamente los cambios se producen por mutación, es decir, un nucleótido puede ser reemplazado por otro, y esto se detecta al obtener la secuencia del gen.
Al origen de nuevas especies se le denomina especiación. Involucra dos procesos: 1] el origen de diferencias genéticas entre poblaciones de una misma especie, y 2] la diferenciación fenotípica entre ellas. Como se mencionó antes, para que ocurra la diversificación entre organismos, es decir, la especiación, es necesario que se originen barreras reproductivas entre las poblaciones. El tipo de especiación se determina de acuerdo con las circunstancias geográficas que originan las barreras reproductivas.
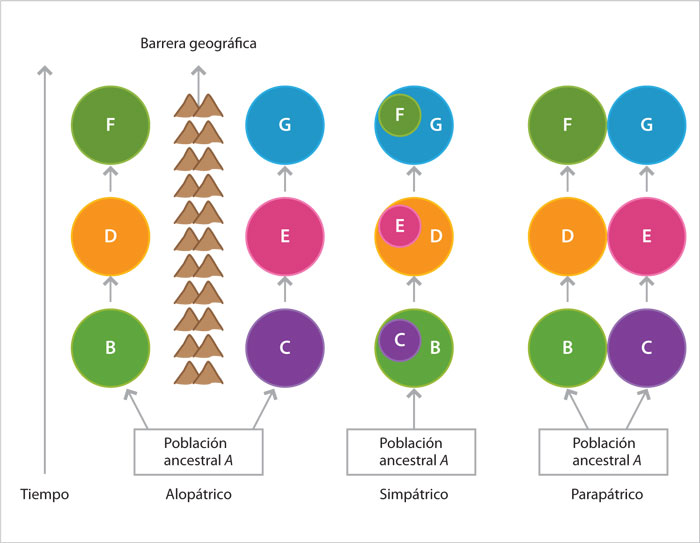
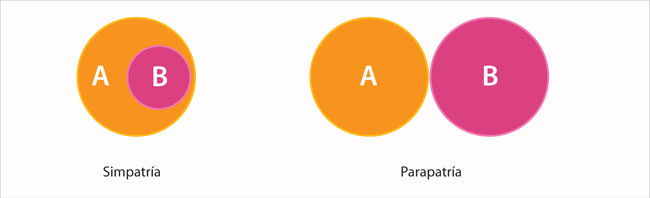
Consideremos que una especie tiene dos poblaciones (A y B), separadas por barreras geográficas (un río, una montaña o un brazo de mar). Si las barreras reproductivas, y por tanto la diferenciación genética y fenotípica, ocurren en esa situación, el proceso de especiación se denomina alopátrico (que significa "en distintos sitios o patrias"). Por el contrario, si la formación de la barrera reproductiva se origina cuando las poblaciones A y B comparten al menos una fracción de hábitat, el proceso se denomina simpátrico ("en el mismo sitio o patria"). Un modelo intermedio a los anteriores, denominado parapátrico (que significa "al lado de"), indica que A y B son contiguas.
El proceso de especiación puede involucrar muchas generaciones de organismos y, por tanto, intervalos de tal magnitud que hacen imposible observar el origen de especies nuevas en tiempo humano. Sin embargo, es posible reconstruir el fenómeno debido a que distintas poblaciones se encuentran en especiación. Para ello se cuenta con las herramientas de la genética molecular, la biogeografía, la ecología y la sistemática.
El siguiente diagrama ilustra de manera esquemática los diferentes modelos de especiación. Las letras hacen referencia a las poblaciones que se originaron, en distintos tiempos, a partir de una ancestral denominada A. Los círculos remiten a la distribución geográfica de una población, y las diferencias entre círculos indican cuándo se ha producido la diferenciación entre poblaciones y el inicio del proceso de especiación.
En el modelo alopátrico la diferenciación ocurre en aislamiento geográfico, producido por una barrera que impide el cruzamiento entre individuos de las dos poblaciones, mientras en el simpátrico una población se diferencia dentro del mismo lugar que ocupa la población original.
En el modelo parapátrico la diferenciación ocurre entre poblaciones vecinas. Puesto que tanto en el modelo simpátrico como en el parapátrico se entrecruzan individuos de las dos poblaciones, es necesario que la selección natural elimine a los híbridos para que se constituya una nueva especie.
El proceso de especiación es un fenómeno común en la naturaleza. Prueba de ello es la gran diversidad de especies que existen en los distintos grupos. Sin embargo, también ocurre el proceso opuesto: la extinción. El registro fósil es una prueba contundente de que las especies no existen por siempre, a pesar de que muchas de ellas permanecieron en el planeta durante millones de años. Hoy día, muchas extinciones han sido provocadas por los humanos, y la desaparición acelerada de las especies actuales podría quizá equipararse con las llamadas extinciones masivas que han ocurrido en la Tierra.
El origen geográfico o alopátrico de nuevas especies significa que las diferencias genéticas y reproductivas entre poblaciones hermanas se dieron cuando ya se habían separado. La especiación alopátrica puede ser resultado de cambios aleatorios en la estructura genética de las poblaciones (véase deriva génica en este mismo tema), o bien, y quizá más frecuentemente, producto de la adaptación de las poblaciones a distintas condiciones ambientales.
Una manera en la que puede ocurrir la especiación alopátrica es la siguiente: dos poblaciones (A y B) de una especie de planta se desarrollan en dos hábitats distintos y separados, de manera que los organismos de una población no se entrecruzan con los de la otra. Las condiciones del hábitat A son de mucha aridez y el hábitat B se encuentra en una zona montañosa con temperaturas muy bajas y una estación de crecimiento muy corta. Para subsistir en la población A es necesario que la selección natural favorezca a los organismos que posean las características para resistir la sequía y elimine a los no resistentes. Por el contrario, en la población B la selección favorecerá a los individuos capaces de resistir las heladas y reproducirse en intervalos de tiempo cortos. Si esto ocurre así, en un sitio A tendremos organismos adaptados a la falta de agua, y en el B a organismos de rápida reproducción y acondicionados a vivir en temperaturas bajas. Este proceso de selección natural divergente entre ambientes produce diferenciación genética y fenotípica entre poblaciones, así como una adaptación local, es decir, poblaciones ajustadas a condiciones ambientales particulares.
En este escenario, la especiación ocurre como una consecuencia de la adaptación local y de los cambios genéticos ocurridos en poblaciones separadas geográficamente. Un ejemplo de ello lo constituye la planta Potentilla glandulosa. Ésta habita en Estados Unidos, desde la costa en San Francisco hasta las montañas de Sierra Nevada. Las poblaciones de la costa, las de media montaña y las de la zona alpina difieren en tamaño, resistencia a las heladas y tiempo de floración. Cuando los individuos de la costa se trasplantan en la zona montañosa no pueden sobrevivir, y viceversa. Este tipo de experimentos se denominan de trasplantes recíprocos. Asimismo, las plantas de la costa florecen desde marzo, pero las de la montaña lo hacen en julio, de manera que no podrían cruzarse, esto se denomina aislamiento reproductivo temporal. Se espera que la especiación ocurra como consecuencia de la adaptación de las poblaciones a distintos factores del ambiente.
En ocasiones un hábitat puede ser muy variable o heterogéneo a pequeña escala. Por ejemplo, si se camina por un bosque, se caerá en cuenta de que existen sitios sombríos, donde no penetra el sol a nivel del suelo debido a las copas de los árboles, y sitios con una alta luminosidad, en claros donde ha caído un árbol. En las selvas existen especies que pueden vivir como juveniles en la umbría del bosque y que se denominan tolerantes a la sombra, pero también hay otras que sólo pueden crecer en los claros con alta radiación lumínica; estas especies se denominan pioneras.
En casi todos los bosques hay especies pioneras y tolerantes. Aunque las segundas son más abundantes, las primeras constituyen una parte importante de los bosques. ¿Por qué existen tantas especies pioneras si unas pocas o una sola podría cumplir con el papel ecológico de colonizar los claros? Es aquí donde la especiación entra en escena. Debido a que la diferenciación de las poblaciones ocurre en el mismo sitio, existe el potencial para entrecruzarse con miembros de otra población en el mismo bosque. Si ocurre la diferenciación, el proceso de especiación será el simpátrico. Este modelo no requiere aislamiento geográfico entre las poblaciones como el alopátrico; por ello, es difícil de explicar cómo ocurre y qué factores facilitan su aparición.
En primera instancia, uno de los factores fundamentales que favorece la diferenciación de las poblaciones en simpatría es la heterogeneidad ambiental a pequeña escala. Un caso sorprendente de diferenciación en simpatría ha ocurrido en un género de moscas de la fruta, Rhagoletis, de la familia Tephritidae. Estas moscas buscan un árbol frutal para copular y las hembras depositan sus huevos en los frutos en desarrollo. Una vez que eclosionan las larvas, consumen la pulpa de la fruta (usualmente especies de la familia Rosaceae: duraznos, cerezas, peras, capulines, ciruelos, chabacanos, tejocotes y manzanos). Cuando llegan a la fase de pupa o crisálida, entran en una etapa de latencia y emergen como adultos en la siguiente primavera, cuando se reproducen para continuar con su ciclo de vida.
En Estados Unidos, las especies de Rhagoletis depositan sus huevecillos en especies del género Crataegus (al cual pertenecen los tejocotes), a las que se denomina plantas huéspedes. Sin embargo, en el siglo XVIII se introdujeron los manzanos de Europa, así que las rosáceas nativas fueron eliminadas de muchos sitios y en su lugar se plantaron manzanos. Las moscas son actualmente una plaga en huertos de manzanos, cerezas y peras. Hoy día también es posible apreciar que existen poblaciones de Rhagoletis adaptadas a usar como plantas huésped especies como el manzano, que antes no existían en esos sitios. Los machos esperan en estos frutos para copular con las hembras y éstas ponen sus huevos en la pulpa. De esta forma, la selección natural ha favorecido que las moscas que crecen en los tejocotes se reproduzcan en tejocotes, y las que crecieron en manzanos, se reproduzcan sólo en manzanos. Este fenómeno se denomina especiación por cambio de huésped y produce también adaptación local. Aunque entre ambas poblaciones existe posibilidad de entrecruzamiento, esto no ocurre, pues entre ellas ya hay una diferencia en sus frecuencias alélicas y porque sus ciclos de vida varían: las moscas de los manzanos se desarrollan en 40 días, mientras las de los tejocotes entre 55 y 60 días. Así, esta diferencia constituye una barrera reproductiva que favorece la divergencia.
Alfred Russel Wallace definió la manera en que la selección natural puede favorecer la especiación al eliminar organismos que son producto del cruzamiento entre poblaciones diferenciadas. Este fenómeno selectivo se conoce como el efecto Wallace o la teoría del refuerzo. En esencia, establece que la selección natural favorece el reconocimiento genético, fenotípico o conductual entre individuos de una misma población para que puedan reproducirse. Aquellos que se entrecrucen con organismos de otra población no tienen posibilidad de dejar descendencia.
En la especiación en simpatría y parapatría ocurre el efecto Wallace para consolidar las barreras reproductivas. La especiación alopátrica sólo requiere un reforzamiento si las poblaciones vuelven a ocupar el mismo hábitat. Considérese una población A adaptada a condiciones de aridez (baja disponibilidad de agua) y una población B resistente a las heladas. Para facilitar la comprensión del fenómeno, supóngase que en cada caso existe un gen que determina la resistencia a la sequía (A) con dos alelos: As Ans, este último alelo dominante; y el gen para la resistencia a las heladas (B), igualmente con dos alelos Bhy Bnh. Si los organismos son diploides como los humanos, tendrán dos copias de cada gen; entonces se verá que los genotipos en cada población serán:
AsAs/BnhBnh para la población A. El gen As determina la resistencia a la sequia.
AnsAns/BhBh para la población B. El gen B determina la resistencia a las heladas.
Los genes Ans y Bnh indican la no resistencia a la sequia y las heladas. Son genotipos mal adaptados.
Si las dos poblaciones se encuentran en simpatría o en parapatría, como se ilustra, entonces los cruzamientos entre individuos pueden producir descendencia con los siguientes genotipos:
Si un individuo de la población A se cruza con otro de la misma población, su descendencia estará adaptada para resistir la sequía.

Es decir, los descendientes no tienen ventajas en ninguno de los ambientes. Por tanto, los individuos que se crucen con otros de distinta población tendrán escasas probabilidades de dejar descendencia, por lo que se espera que la selección natural favorezca las características que faciliten el reconocimiento de los individuos de la misma población.