
![]()
![]()
Podemos distinguir tres tipos de variación: fenotípica, genotípica y geográfica. La variación fenotípica se refiere a las variantes que existen en la morfología, fisiología, bioquímica y comportamiento de los organismos. La variación genética indica las diferencias en las secuencias del ADN en los individuos de una población. Como el fenotipo es resultado de los genes, del ambiente y de su interacción, también se encuentran distintas expresiones fenotípicas que dependen del área donde se desarrollan los organismos, lo que se ha denominado variación geográfica.
En biología evolutiva se hace referencia a la variación genética en dos niveles distintos: la variación individual y la variación poblacional. La variación individual es generada por dos procesos moleculares: la mutación y la recombinación genética; el primero genera nueva variación, mientras que el segundo aporta una cantidad ilimitada de nuevas combinaciones genéticas. La variación poblacional son los cambios en las frecuencias de las variaciones genotípicas y fenotípicas a nivel de la población. Estos cambios son consecuencia de procesos evolutivos como la selección natural, la migración y la deriva génica, entre otros, los cuales son causa de la transformación de las poblaciones y, por tanto, de las especies.
La variación fenotípica entre individuos puede entenderse como la diferencia en uno o más caracteres que ha resultado de la interacción genoma-ambiente. Un gran número de características de los organismos muestra diferencias entre poblaciones de una especie, y son de distinta naturaleza. Si se trata de diferencias cualitativas, como la presencia o ausencia de un rasgo, por ejemplo de bandas en la concha de los caracoles, se denominan caracteres discretos, justo como los que Mendel analizó en los chícharos. Algunas características de los organismos, como la masa corporal, la estatura o la longitud, poseen variación continua, es decir, existe un número muy grande de valores fenotípicos en una población, por lo que es difícil distinguir dos fenotipos similares entre sí.
La variación en la mayoría de los caracteres de los organismos forma un continuo fenotípico, más que clases discretas. Los caracteres, como la masa corporal, por ejemplo, que se distribuyen estadísticamente de manera continua, se denominan métricos. Los que muestran un número elevado de fenotipos en las poblaciones, pero que se presentan de manera discreta, se denominan merísticos. Ejemplos de éstos son el número de huevos que pone un ave o el número de frutos o semillas de una planta. Existen rasgos para los cuales sólo conocemos dos estados: presencia o ausencia. Éstos son los caracteres umbral, es decir, existe un valor fenotípico a partir del cual se posee la condición. Ejemplo de éstos son ciertas condiciones metabólicas como la diabetes. Es muy probable que los individuos de la población varíen de manera continua en su propensión a esta enfermedad.
El papel que juega la variación fenotípica en el proceso de selección natural puede explicarse con dos ejemplos sencillos:
1. El caracol terrestre Cepaea nemoralis presenta variación en la coloración y en el número de rayas de su concha. Existen caracoles con concha amarilla, rosa o café. Además, pueden no tener rayas o tener de una a cinco. Este caracol habita en bosques donde el color del suelo es uniforme y oscuro, así como en pastizales. ¿Cuál es la razón para que existan tantas formas de la misma especie? Una posibilidad es que la coloración de la concha ofrezca camuflaje. Los caracoles son alimento de los tordos (Turdus philomelos), los cuales rompen las conchas con piedras; por tanto, puede saberse qué tipo de caracoles son más comidos por los tordos en cierto sitio.
Por medio de experimentos sencillos los biólogos encontraron que la coloración de la concha determina la probabilidad de vivir o morir para un caracol. En un ambiente delimitado se contaron los caracoles vivos y muertos. Ya que los tordos dejan la concha, es fácil determinar cuáles sirvieron de alimento. Los resultados confirmaron la hipótesis de que la coloración ofrece camuflaje y por ello propicia la supervivencia: en ambientes oscuros como el bosque, los caracoles de color café y sin rayas son más abundantes; pero en el ambiente claro de los pastizales proliferan los de color amarillo y con rayas.
2. En los humanos, los eritrocitos transportan el oxígeno a todos los órganos y tejidos, ya que poseen hemoglobina. Ésta es una proteína de cuatro unidades: dos denominadas beta y dos alfa. Los individuos con anemia falciforme presentan eritrocitos en forma de hoz. El alelo normal se representa como HbA (Hb por hemoglobina), mientras que la hemoglobina alterada es HbS. Los individuos sanos tienen un genotipo HbAHbA, mientras quienes tienen anemia son HbS HbS y tienen una probabilidad de sobrevivir muy baja; los heterocigotos HbAHbS son ligeramente anémicos. La diferencia entre los alelos HbA y HbS reside en un cambio de aminoácido en la posición 6 de la cadena beta de la hemoglobina: la más común tiene ácido glutámico y la otra hemoglobina tiene valina. Esta sustitución provoca la cristalización de la hemoglobina cuando hay baja presión de oxígeno y entonces el eritrocito se deforma y desintegra.
Sería de esperarse que, dado que los individuos con homocigotos HbSHbS por lo general mueren, en el proceso de selección natural desapareciera por completo el gen nocivo, y así ocurre en la mayoría de las poblaciones humanas, excepto en algunas regiones de África, donde hay incidencia de malaria o paludismo (enfermedad causada por un protista llamado Plasmodium, transmitido por mosquitos). Los individuos heterocigotos (HbAHbS) son resistentes a la malaria, pero no los homocigotos normales (HbAHbA). En este caso se dice que hay una ventaja del heterócigo; los homocigotos normales HbAHbA presentan mayor mortalidad ante el paludismo que los heterocigotos HbAHbS debido a que el parásito causante de la enfermedad se desarrolla dentro de los eritrocitos y éstos, en los heterocigotos, se destruyen con más facilidad que los eritrocitos normales, por lo que el desarrollo del parásito no prospera. Como consecuencia, los heterocigotos resistentes a la malaria tienen mayor éxito en la supervivencia y reproducción. En este caso la selección natural mantiene la variabilidad genética, pues los heterocigotos tienen ambos alelos.
Por variación genética se entienden las diferencias en el material hereditario de los individuos de una población, ya sea en los cromosomas o en las secuencias de ADN. Ésta explica, por ejemplo, algunas diferencias entre las personas, como el color de ojos y el grupo sanguíneo. La variación genética puede jugar un papel importante en el riesgo de que una persona padezca alguna enfermedad particular; sin embargo, es importante considerar que muchas de las enfermedades son resultado de una interrelación compleja entre múltiples genes y el ambiente, tales como la diabetes o las enfermedades del corazón. En la biología evolutiva, variación genética también se refiere a los cambios de las frecuencias génicas en la estructura de una población, cambios que son resultado de procesos evolutivos como la selección natural, la migración y la deriva genética.
En las especies diploides con reproducción sexual la diversidad genética total de la población está representada por todos los individuos, machos y hembras reproductivamente maduros, que tienen la posibilidad de contribuir, con descendencia, a la generación siguiente. La variabilidad genética de una población se denomina pool génico o acervo de genes. Las causas que afectan la probabilidad de que los individuos hereden sus genes a la generación siguiente afectan, a su vez, la composición genética y por tanto la evolución de la población.
En los organismos diploides, como los humanos, todas las células, con excepción de los gametos, poseen dos copias del genoma completo de la especie; una es heredada de la madre y otra del padre. En los seres humanos el genoma completo está distribuido en 23 pares de cromosomas: 22 pares de autosomas y un par de cromosomas sexuales que determinan el sexo del individuo. Por tanto, cada gen contenido en los autosomas posee dos copias, una en cada uno de los cromosomas homólogos. Si las dos copias del gen son similares, el individuo es homocigoto (Ai = Ai) y si son distintas, es heterocigoto (Ai ≠ Aj). Aun cuando cada individuo de la especie posee dos copias de cada gen, en la población en su conjunto pueden existir varios alelos para el mismo gen. Un ejemplo de esto son los alelos que determinan los tipos sanguíneos ABO en humanos: existen tres alelos, IA, IB e i. Cada ser humano tiene dos de estos alelos, pero la población posee tres. A pesar de existir seis genotipos distintos, sólo cuatro fenotipos se observan en las poblaciones humanas debido a que los alelos IA e IB son codominantes entre sí y dominan sobre el recesivo i (véase cuadro 2).

Si la variación en las poblaciones es un requisito indispensable para la evolución, una pregunta obligada es: ¿cuánta variación genética existe en las poblaciones naturales? Un problema inicial para medir la variación genética en las poblaciones es que, como se dijo anteriormente, la relación entre genotipo y fenotipo no es directa y por ello no podemos cuantificar la variación genética sólo con medir la variación fenotípica.
A principios del siglo XX se redescubrieron las leyes de Mendel. Esto significa que con anterioridad no se conocía la base genética de muchas variantes fenotípicas. Sin embargo, este redescubrimiento brindó la posibilidad de cuantificar la variación. Durante los primeros años del siglo XX nació la genética de las poblaciones, disciplina que se ocupa de la descripción de las poblaciones desde el punto de vista genético, la diversidad de genotipos y las variantes alélicas de cada gen, así como de las causas que producen cambios en dicha composición.
La población puede ser descrita desde el punto de vista genético debido a que los individuos que la componen son los poseedores de los genes que serán transmitidos a la generación siguiente y, por tanto, sólo la población es susceptible de evolucionar en su composición fenotípica y genética. Puede describirse genéticamente a la población de dos maneras: 1] a través de la frecuencia o proporción de cada genotipo (frecuencias genotípicas), y 2] por medio de la frecuencia o proporción de cada gen o alelo (frecuencias génicas o alélicas).
Para ilustrar la descripción genética puede suponerse lo siguiente:
1. Se trata de una especie diploide con reproducción sexual.
2. Existe un número grande de individuos (N).
3. Se obtendrán los genotipos de un locus (A), que posee dos alelos (A1 y A2) en la población (y por tanto hay tres genotipos posibles: A1A1, A1A2 y A2A2)
4. Existen n11 , n12 y n22 individuos de cada genotipo.
5. Ya que se conoce qué alelos porta cada genotipo, es posible obtener la frecuencia de cada uno (A1 o A2).
En los cuadros siguientes (3 y 4), puede apreciarse cómo se hace el cálculo para obtener las proporciones genotípicas y alélicas. La suma de las frecuencias genotípicas en la población es 1.

De manera similar, debido a que se conocen los alelos de cada genotipo, es posible calcular la frecuencia de cada gen. Por ejemplo, los individuos homocigotos A1A1 llevan dos copias del mismo alelo (ya que son diploides), mientras el heterocigoto lleva una copia del alelo A1 y una copia del alelo A2. El número total de alelos en la población será igual a dos veces el tamaño de la población de individuos o 2N. Véase el cálculo:
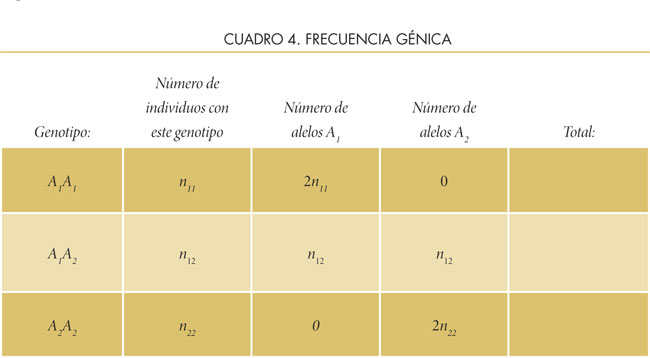
1. La frecuencia del alelo A1será:
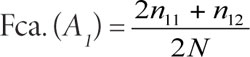
2. La frecuencia del alelo A2será:
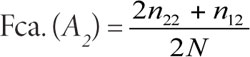
Si se representa la frecuencia del alelo A1 como p y la del alelo A2 como q, entonces pueden simplificarse las expresiones de la siguiente manera:
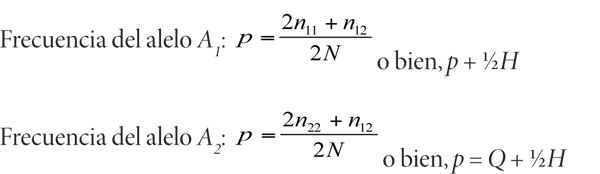
Se tienen ahora dos modos para describir genéticamente a la población a través de las frecuencias genotípicas y alélicas. Si se considera a la evolución en una escala de tiempo pequeña y se define como el cambio en la frecuencia de los genes y de los genotipos, a través del tiempo o entre generaciones puede conocerse: a] el proceso que produce el cambio en la población, y b] la velocidad a la que lo produce. Así, la evolución de la población estará determinada por el cambio en las proporciones de los fenotipos y genotipos.
La expresión de algunos genes es afectada por el ambiente donde se desarrolla el individuo. Por ejemplo, está el caso de la planta Potentilla glandulosa, pariente de los rosales, que crece desde la costa del Pacífico hasta la zona alpina de la Sierra Nevada, en el estado de California, Estados Unidos. A lo largo de esta distribución geográfica —desde la costa hasta la cima de la sierra—, los individuos muestran diferencias fenotípicas entre localidades: en la zona alpina son muy pequeñas, en la costa son altas, y en la zona de media montaña, esbeltas. A este tipo de variación se le denomina variación geográfica.
Se realizó un experimento para determinar si las diferencias fenotípicas entre las plantas de las diferentes altitudes se deben a los genes y si son adaptativas en cada ambiente local (altitud). Se eligió un número de plantas de dos altitudes (media montaña y zona alpina) y se realizaron cruzas entre ellas para producir F2. La hipótesis de este experimento es la siguiente: si existen diferencias entre las poblaciones, las cruzas producirán individuos genéticamente intermedios (F1), y posteriormente con la cruza de los F1 se segregará toda la variación de la especie; es decir, se producirán genotipos intermedios o con el genotipo parecido a cualquiera de los progenitores (media montaña o zona alpina).
Una vez obtenida la F2 se efectuó su clonación (es decir, se produjeron individuos idénticos) y se sembró una copia de cada genotipo en tres sitios localizados a diferentes altitudes. Después de varios años se analizó la supervivencia, la edad a la que se producía la floración y el tamaño que alcanzaban los individuos. Los resultados indicaron que los fenotipos variaban ampliamente entre localidades (altitudes) y que mostraban diferentes probabilidades de sobrevivir en distintos sitios: en la alta montaña las plantas de tamaño pequeño, con hojas pequeñas y coriáceas y de floración temprana eran las supervivientes (debido al riesgo de las heladas).
Los resultados del experimento anterior indican que un mismo genotipo (clonado y puesto en distintos ambientes) puede tener distintas expresiones fenotípicas cuando interacciona con su entorno. Además, muestran que los distintos genotipos pueden "reaccionar" de diferente manera a la variación en el medio. Al conjunto de fenotipos que un genotipo puede expresar en distintos ambientes se le denomina norma de reacción. Por supuesto, debido a que en teoría puede existir un número infinito de ambientes, se estudian sólo partes de la norma de reacción de un genotipo (por ejemplo, tamaño o reproducción) y sólo ciertos factores ambientales.
El concepto de norma de reacción fue propuesto en 1909 por el zoólogo alemán Richard Woltereck (1877-1944) a partir de sus estudios con cepas morfológicamente distintas de "pulgas de agua", pertenecientes a los géneros Daphnia y Hyalodaphnia. Estos organismos se reproducen por partenogénesis, de modo que cultivándolos por muchas generaciones, Woltereck pudo obtener líneas puras. Realizó experimentos en los que puso a las diversas líneas en ambientes con contenido variado de nutrimentos (bajo, medio y alto) y midió caracteres métricos, como el tamaño de la cabeza en relación con el tamaño del cuerpo. Sus resultados mostraron que todas las líneas o genotipos tenían normas de reacción muy distintas en otros ambientes. Es decir, aun cuando eran genotípicamente idénticos, colocados en distintos ambientes eran fenotípicamente diferentes.
Los experimentos de Woltereck son muy importantes porque indican que los fenotipos son el resultado de los genes, el ambiente y la interacción entre ellos; es decir, no todos los organismos responden igual a la diferencia en las condiciones ambientales. Esto ha dado origen a la investigación sobre la plasticidad fenotípica, su valor adaptativo y su base genética. La plasticidad fenotípica es la producción de fenotipos alternativos a partir de un mismo genotipo expuesto a distintas condiciones ambientales. Por tanto, a los caracteres que varían conforme se altera un factor ambiental se les denomina caracteres plásticos o que muestran plasticidad fenotípica (genotipos A y B en la figura de Daphnia). Sin embargo, existen caracteres que no varían con el ambiente o que muestran normas de reacción estables (genotipo C en la figura de Daphnia). Estas normas de reacción estables podrían ser el producto de sistemas de amortiguamiento ambiental que se manifiestan durante el desarrollo del organismo, fenómeno que recibe el nombre de canalización. Por ejemplo, existen genotipos altamente canalizados como los tetrápodos (anfibios, reptiles, aves y mamíferos): estos organismos tienen cuatro extremidades que se forman temprano en el desarrollo embrionario; no existe variación en este carácter, por lo que se dice que es un genotipo altamente canalizado. Por el contrario, en Drosophila se observan cambios fenotípicos si las larvas se desarrollan en diferentes temperaturas.