
![]()
![]()
El desciframiento de la configuración molecular del gen inmediatamente planteó un mecanismo detallado para la duplicación y trasmisión de los caracteres hereditarios de una generación a la siguiente. La estructura en doble hélice y la secuencia de bases a lo largo de la cadena permitió concebir un mecanismo preciso de copiado fiel de la información.
Asimismo, de manera también casi inmediata, la secuencia de bases sugirió una relación entre la información contenida en bases nitrogenadas y aquella derivada en forma de proteínas, moléculas ubicuas y abundantes en los seres vivos. En esta sección se plantea el papel del gen en la herencia y en la expresión de esa información en la célula.

El modelo de Watson y Crick tuvo un gran impacto en el desarrollo de la biología molecular al señalar que la información genética estaba contenida en la secuencia de las bases nitrogenadas. El flujo de información genética que liga la información contenida en el ADN, el paso a la molécula intermedia o ARN y la formación de una proteína a partir de esa información fueron descritos como el dogma central de la biología molecular. De acuerdo con él, existen tres flujos básicos de información genética. El primero se refiere a la transferencia de la información genética de una molécula de ADN a otra, durante el proceso de replicación que se efectúa en la fase S del ciclo celular, lo que garantiza su permanencia a través de las generaciones. El segundo se efectúa cuando el ADN que no forma directamente proteínas se sirve de una molécula de ARN como intermediario. La molécula de ARN mensajero se forma en respuesta a una determinada secuencia de ADN; el proceso se denomina transcripción. Las moléculas de ARN mensajero serán usadas por los ribosomas como moldes para dirigir la síntesis de proteínas durante el proceso de traducción, que constituye el tercer camino que sigue la información genética.
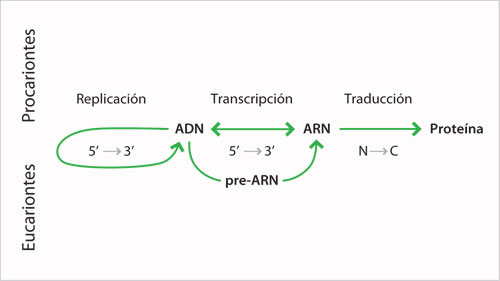
A estas tres vías de flujo de información se han agregado otras: el descubrimiento de los retrovirus, que tienen su información genética codificada en ARN y que poseen una enzima denominada retrotranscriptasa, transfieren la información del ARN a ADN durante el ciclo infectivo para después seguir los caminos regulares de flujo, cuando el ADN viral se incorpore al genoma de la célula huésped; además, el ARN de estos virus se replica para formar nuevas moléculas de ARN que pasarán a las nuevas partículas virales. Por otro lado, se integran en el dogma los genes para la síntesis de ARN ribosómico y ARN de transferencia y los flujos de información que se llevan a cabo en la mitocondria y en el cloroplasto.
La molécula de ADN presenta tres niveles estructurales: la estructura primaria, que se refiere a la secuencia de nucleótidos; la estructura secundaria o doble hélice, y la estructura terciaria, representada por un enrollamiento posterior que permite a una enorme molécula ser empaquetada para caber en el pequeño espacio de una célula procariota (460 millones de bases en el caso de E. coli) o en el núcleo de los organismos eucariontes, como los seres humanos, con los 6 400 millones de pares de bases que se hallan en una célula diploide.
El material genético de las bacterias o nucleoide se encuentra disperso en el citoplasma formando grupos de ADN perfectamente visibles bajo el microscopio electrónico. En la interfase del ciclo celular de las células eucariontes, el ADN y sus proteínas asociadas reciben el nombre de cromatina, que ocupa casi todo el espacio nuclear, pero en territorios bien definidos. Los cromosomas propiamente dicho sólo se hallan en la mitosis, especialmente en la metafase, cuando la cromatina se encuentra más condensada.
La cromatina en su estructura más simple está constituida por la hélice de ADN; en un segundo nivel, la doble hélice y las proteínas histonas forman unidades denominadas nucleosomas. Cada nucleosoma está constituido por dos vueltas de ADN alrededor de un octámero de histonas y una histona adicional que mantiene la estructura en su sitio. Todo el conjunto (ADN y proteínas) tiene el aspecto de un collar de cuentas.
Un tercer nivel de complejidad se presenta al plegarse "el collar de cuentas" de manera apretada alrededor de un eje imaginario; en el siguiente nivel, el solenoide formado de esta manera, se pliega nuevamente y cada plegamiento se ancla a un andamiaje nuclear constituido por fibrillas de proteína. Este nivel es el que se presenta comúnmente en el núcleo en interfase. Cuando el núcleo se divide, la cromatina se condensa aún más para constituir el cromosoma metafásico que es el que se aprecia con el microscopio de luz.
La compactación de la cromatina cambia constantemente adaptándose a los periodos de replicación y transcripción; durante estos periodos, la cromatina tiene que volverse laxa para permitir la entrada de los complejos enzimáticos de ADN polimerasa y ARN polimerasa. De hecho, ciertas zonas de la cromatina permanecen muy condensadas durante la interfase y se caracterizan por estar formadas por repeticiones de secuencias que no se transcriben.

Los cromosomas metafásicos están formados por dos cromátidas exactamente iguales, pues pasaron antes por la fase S del ciclo celular, en la que el ADN se duplicó. Las dos cromátidas se encuentran unidas por una constricción llamada centrómero, que tiene una posición definida en cada uno de los cromosomas. Los centrómeros son esenciales para que se efectúe la mitosis, pues al final de la profase se agregan en este sitio proteínas que forman el cinetocoro, al cual se fijan los microtúbulos del huso mitótico.
Los telómeros constituyen otra parte fundamental del cromosoma; se encuentran en la parte terminal de éste y tienen como función primordial el mantenimiento de la estructura: si los telómeros están ausentes, los extremos de los cromosomas tienden a fusionarse; además, son importantes porque participan en la formación de la estructura tridimensional del núcleo y en el apareamiento de los cromosomas. El ADN de la región telomérica de los cromosomas humanos consta de cientos de repeticiones del motivo 5'-TTAGGG-3'.
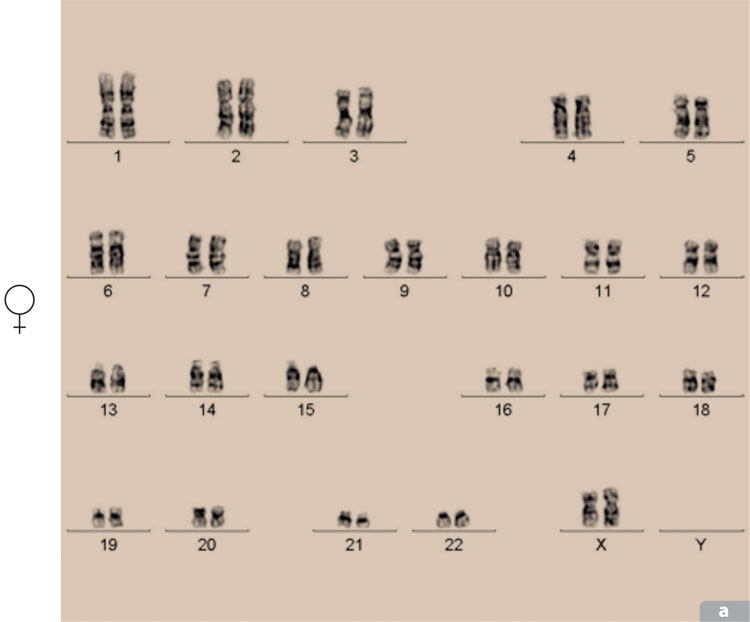
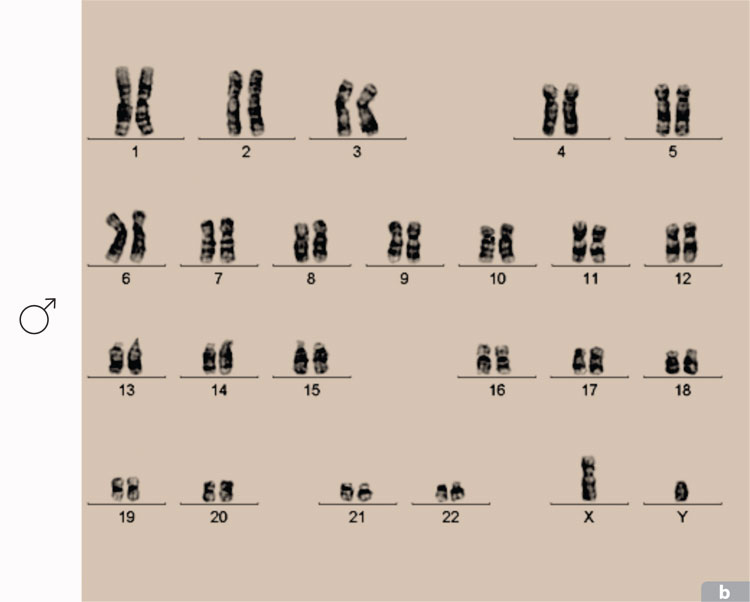
Todos los cromosomas de un organismo pueden representarse por medio de un cariotipo que ordena los cromosomas por su tamaño, por la posición de los centrómeros y por el bandeo obtenido por ciertas tinciones. Por ejemplo, los cromosomas teñidos con el colorante Giemsa despliegan estriaciones denominadas bandas G que permiten identificarlos y analizarlos para reconocer cualquier tipo de alteración, como translocaciones o deleciones. En la actualidad, además del análisis de las fotografías de los cromosomas, se usa la computadora para hacer un análisis más acucioso y la tinción de cromosomas con colorantes fluorescentes.
El término genoma describe el total de la información genética que se encuentra en una especie y en cada una de sus células; en realidad el genoma comprende el ADN del núcleo y el de la mitocondria (y el del cloroplasto en el caso de plantas y otros grupos que presentan este organelo).
El Proyecto Genoma Humano fue concebido para determinar la secuencia de los nucleótidos que constituyen el ADN del núcleo, pues la secuencia del ADN mitocondrial ya se conocía. Desde la mitad de la década de 1970 se iniciaron los trabajos para secuenciar tramos de ADN, cuya primera fase terminó en 2001, con la publicación de la primera versión de la secuencia en las revistas científicas Nature y Science.
El análisis de la secuencia parcial (aproximadamente 2 600 millones de bases) reveló la composición del genoma humano: sólo una pequeña parte se transcribe a ARN mensajero; el resto del ADN está formado por intrones y fragmentos de genes, pseudogenes que son copias no funcionales de algunos genes; secuencias moderadas y altamente repetidas; microsatélites y transposones (secuencias capaces de insertarse en un nuevo sitio dentro del genoma).
De acuerdo con lo anterior, únicamente una pequeña parte del ADN de los eucariontes sigue el esquema del dogma central, al transcribir su información para formar proteínas. Además, según las necesidades de cada célula en particular, sólo algunas regiones del genoma, separadas por largos tramos de ADN no codificante, se comportan como unidades de transcripción. Estas unidades o genes se encuentran dispersos en el ADN de una manera irregular.
La presencia de grandes espacios que se encuentran entre las regiones codificantes de los genes, impiden que el ARN mensajero se forme directamente; las enzimas que participan en el copiado de la información sintetizan una primera versión de ARN que copia todo el tramo de ADN, las regiones codificantes y las no codificantes (transcrito primario), el cual se modifica posteriormente al eliminar las regiones no codificantes del transcrito primario.
Como se mencionó anteriormente, a diferencia de los genes de los procariontes, los genes de los eucariontes no se corresponden directamente con el ARN mensajero, debido a que entre los exones (regiones que codifican para el péptido) se encuentran los intrones, que no se relacionan con la secuencia de los aminoácidos.
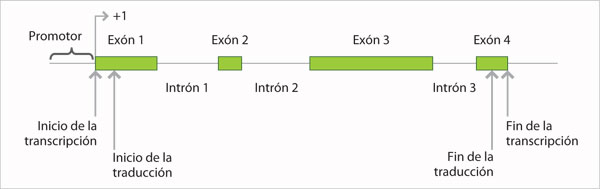
La estructura típica de un gen de eucarionte con tres exones consiste de:
• Una región promotora que es indispensable para que se inicie la transcripción. Aquí se encuentra la caja TATA, que es el sitio de unión de la enzima ARN polimerasa que cataliza la polimerización de ARN a partir de un molde de ADN. Se encuentran también otras regiones que intervienen en la regulación de la transcripción.
• El exón 1 presenta un sitio de inicio de la transcripción, que es importante porque aquí se agrega un "capuchón" de GTP metilado, que impedirá la destrucción del ARN.
• Entre el sitio de inicio de la transcripción y el sitio de inicio de la traducción se encuentra una región que no se traducirá y que recibe el nombre de región no traducida 5' (5'UTR, 5' untranslated region). Esta zona determina la velocidad de inicio de transcripción.
• El sitio de inicio de la traducción está marcado por el codón ATG, que en el ARN corresponde a AUG o codón de inicio de la traducción.
• El resto del exón contiene los pares de bases que codificarán para los primeros aminoácidos del péptido.
• El intrón 1 es seguido por el exón 2 y el intrón 2. El exón 3 contiene los últimos pares de bases que codificarán para el resto de los aminoácidos.
• Además, este exón presenta dos zonas importantes: el codón de terminación que puede ser TAA que se convertirá en UAA en el ARN mensajero y la secuencia AATAAA, a la cual se agregará una cola de poli A que tiene la misma función que el capuchón inicial.
• Entre el codón de terminación y la secuencia AATAAA se encuentra la región no traducida 3' (3'UTR, 3' untranslated region).
• La transcripción continúa más allá de la región no traducida.
La expresión de los genes ocurre en dos partes:
• La transcripción genera una hebra de ARN mensajero idéntica en secuencia a la hebra de ADN orientada en dirección 5' a 3'.
• Sólo una de las hebras del ADN es transcrita a ARNm. La hebra que dirige la formación de ARN mensajero mediante el apareamiento de las bases se denomina hebra molde o hebra antisentido y tiene una dirección de 3' a 5'.
• La otra hebra de ADN posee la misma secuencia del ARNm, aunque en lugar de uracilo tiene timina. Esta hebra se denomina hebra codificante y al igual que el ARNm corre de 5' a 3'.
• La traducción convierte una parte de la secuencia de ARNm en una secuencia de aminoácidos. La región codificante consiste de una serie de codones, cada uno de los cuales representa un aminoácido.
Durante las décadas de 1960 y 1970, los análisis de los genes bacterianos mostraron que existía una correspondencia uno a uno entre los codones del ARNm y del ADN. Por esta razón, el ARNm que se formaba sobre la hebra codificante de ADN no necesitaba ninguna modificación posterior. Sin embargo, en la década de 1970 se descubrió que en los eucariontes no existía relación colineal entre el ARNm y el ADN. Se encontró que los genes eucariontes tenían tramos codificantes separados por tramos (en ocasiones muy largos) de regiones no codificantes; por lo tanto, el proceso de transcripción de los eucariontes debía ser diferente del de los procariontes.
Al inicio del proceso, toda la secuencia del gen se transcribe a ARN. Posteriormente, los intrones se quitan y los exones quedan unidos. Este proceso se denomina splicing, que es característico de los genes eucariontes, aunque algunos procariontes también lo tienen. El ARN tiene otras modificaciones adicionales, por ejemplo, se agrega la caperuza de metil guanosín en la región 5' y la cola de poli A en la región 3', que evita la degradación del ARNm por enzimas de tipo exonucleasas.
En los procariontes, debido a la ausencia de una envoltura nuclear, la transcripción y la traducción ocurren simultáneamente, de tal manera que conforme se va sintetizando el ARNm, los ribosomas se adhieren a él para efectuar la síntesis de la proteína. Pero en los eucariontes, después de formado el ARNm, éste sale del núcleo y se dirige hacia el citosol, donde se encuentra con las subunidades pequeña y grande de los ribosomas. Los ribosomas constituidos por ARN ribosómico y diversas proteínas se unen a la región 5' del ARNm y se mueven hacia la región 3' traduciendo a su paso los codones del mensajero. El proceso de traducción consta de tres partes: iniciación, elongación y terminación.
Durante la primera fase se forma un complejo de iniciación integrado por las subunidades grande y pequeña del ribosoma que se unen en el sitio del codón de inicio del ARNm, y por el ARN de transferencia que se encuentra unido por medio de su anticodón al codón de inicio del ARNm; este ARN de transferencia lleva el aminoácido metionina y se coloca en el sitio P (por la palabra péptido) del ribosoma.

Después de que se ha integrado el complejo de iniciación, el ribosoma se desliza hacia el extremo 3' del ARNm. En cada deslizamiento, el ribosoma se desplaza un codón permitiendo que el péptido en crecimiento quede colocado en la posición P, mientras el sitio A (por la palabra aminoácido) es ocupado por un RNA de transferencia cuyo anticodón debe corresponder con el codón colocado en esta posición. Cada vez que un ARN de transferencia se coloca en el sitio A, su aminoácido se une al péptido situado en el sitio P, mediante un enlace peptídico catalizado por una molécula de ARN ribosómico de la subunidad grande del ribosoma (ribozima). En seguida, el ARN de transferencia del sitio P se desprende del ribosoma, sale de él a través del sitio E (del inglés exit: salida) y se dirige al citosol para cargarse nuevamente con el aminoácido que le corresponde.
Al llegar al codón de terminación, ciertas proteínas denominadas factores de terminación se unen a este codón provocando la liberación del péptido. Cuando los sitios P y A del ribosoma no tienen ya ARN de transferencia, las dos subunidades del ribosoma se separan.
La traducción en procariontes y eucariontes se lleva a cabo simultáneamente por muchos ribosomas configurando una estructura llamada polirribosoma que está formada por los ribosomas unidos al ARNm. Cada ribosoma se une al extremo 5' del ARNm y se mueve hacia la región 3', de tal modo que conforme se desplaza, el péptido que sintetiza va creciendo progresivamente.
Cuando las células se dividen, las células resultantes tienen la misma información genética. Sin embargo, en ciertas ocasiones ocurre una mutación, es decir, un cambio permanente en el material genético que pasa de una generación celular a la siguiente (mutación somática); si la mutación afecta a las células reproductoras, el cambio genético puede pasar de los padres a la progenie (mutación gamética).
Las mutaciones explican la variación alélica que se ejemplificó al analizar las leyes de Mendel con la presencia de plantas de chícharo con fenotipo de tallos altos y bajos o de semillas rugosas y lisas. No siempre se aprecian los cambios fenotípicos producto de las mutaciones; por ejemplo, ciertos cambios genéticos se apreciarán sólo si determinadas sustancias están presentes en las células. La enzima glucosa 6 fosfatodeshidrogenasa, que normalmente está presente en muchas de las células del organismo humano, debido a una mutación puede presentarse en niveles extremadamente bajos. Su presencia sólo se apreciará por la ruptura de los eritrocitos de un individuo que tenga la mutación y que esté bajo los efectos de la primaquina, que es un medicamento contra la malaria.
Las mutaciones pueden ser benéficas, nocivas o neutras. Los efectos positivos de la mutación se reflejan en los cambios que se presentan en las poblaciones y que les permiten adaptarse a las modificaciones del medio. Este tipo de mutación es la base de la evolución.
Las mutaciones nocivas impiden que los genes desempeñen su función de manera adecuada. Puesto que las mutaciones pueden ser extremadamente dañinas al punto de poner en peligro la integridad del material hereditario, los organismos han desarrollado diversos mecanismos para reparar los errores. Las mutaciones pueden dividirse en 1] mutaciones puntuales o mutaciones por sustitución de bases, 2] mutaciones cromosómicas y 3] mutaciones genómicas. Las del primer tipo, en las que se reemplaza un nucleótido por otro, se dividen en transiciones, cuando una pirimidina se cambia por otra pirimidina (C por T) o una purina por otra purina (G por A) y en transversiones, que se caracterizan porque un par de bases simplemente cambian su posición de manera transversal. Además de éstas se pueden agregar o quitar cortas secuencias de nucleótidos denominándose inserciones o deleciones, respectivamente. Las mutaciones puntuales tienen diversos efectos sobre el tipo de aminoácidos que forman la cadena de polipéptidos.
Las mutaciones silenciosas no alteran la secuencia de los aminoácidos, a pesar de que la secuencia de bases está modificada; debido a la existencia de un código genético redundante pero no ambiguo (código degenerado), las mutaciones pueden afectar a ciertas bases del codón, sin que el aminoácido cambie.
En las mutaciones cambio de sentido, una mutación provoca el cambio de un aminoácido por otro. Así, la cadena b de la hemoglobina que en su posición 6 normalmente tiene ácido glutámico, en la mutación que causa la anemia falciforme aparece una valina; dado que hay un cambio de un aminoácido polar por un hidrofóbico, la conformación de la proteína cambia, de tal modo que provoca el aglutinamiento de las moléculas de hemoglobina, con la consecuente deformación de los eritrocitos.
Las mutaciones sin sentido involucran la inserción de un codón de terminación en lugar de un codón normal. Por esta causa, la síntesis del polipéptido se trunca, debido a la presencia inapropiada del terminador.
No todas las mutaciones puntuales se refieren a sustituciones de bases. Las inserciones y deleciones, al insertar o quitar una base en cualquier punto de la secuencia del gen, cambian totalmente la disposición de los codones y, por lo tanto, el tipo de aminoácidos que se integrará al polipéptido. Estas mutaciones, que pueden ser muy graves, se denominan de cambio de fase o de cambio de marco de lectura.
Las mutaciones cromosómicas alteran la estructura de un cromosoma y por lo tanto afectan a más de un gen. La modificación de la estructura cromosómica se realiza de dos maneras: el material cromosómico puede aumentar o disminuir de manera significativa dentro de un mismo cromosoma (duplicaciones y deleciones); o bien, el material genético puede reorganizarse sin modificar la cantidad total de material genético (inversiones y translocaciones).
La deleción o deficiencia se realiza cuando un segmento del cromosoma desaparece y, por consiguiente, varios genes son los afectados; mientras la duplicación ocurre cuando una sección del cromosoma se repite a lo largo del mismo. Las consecuencias de las deleciones dependen del tamaño del fragmento suprimido y si los genes afectados son necesarios para el desarrollo del organismo. En los humanos, la enfermedad cri-du-chat (maullido de gato), caracterizada por retraso mental, anomalías faciales y sonidos que recuerdan el maullido de un gato, es causada por la supresión de un segmento del brazo corto del cromosoma número 5.
Las duplicaciones o segmentos repetidos de un cromosoma tienden a ser menos nocivas que las deficiencias; sin embargo, existen algunos problemas relacionados con pequeñas duplicaciones como la enfermedad de Charcot-MarieTooth, neuropatía caracterizada por adormecimiento de manos y pies.
Además de su innegable interés médico, las duplicaciones tienen un importante papel en la evolución de los genes y, por lo tanto, de las proteínas. La copia de un gen puede mutar y adquirir nuevas funciones a través del tiempo. Los genes relacionados de esta manera tienen una secuencia de aminoácidos muy similar, como en el caso de los que codifican para tripsina y pepsina o los que dan origen a hemoglobina y mioglobina. La existencia de las familias de genes también puede explicarse por el mismo fenómeno: grupos contiguos de genes con pequeñas mutaciones, cuyos productos efectúan la misma función, por ejemplo, los genes que codifican para los diferentes tipos de hemoglobinas humanas.
Las inversiones son otro tipo de mutación cromosómica en las que tramos de ADN giran
180º. Este tipo de mutación no implica pérdida de información sino una simple reorganización de la misma. Para entender las consecuencias de las inversiones se debe recordar que durante la meiosis, las cromátidas de cromosomas homólogos normalmente se entrecruzan para promover una gran cantidad de diversidad genética. Si un individuo porta una copia de un cromosoma normal y un homólogo que ha sufrido una inversión, habrá una gran probabilidad de producir gametos anormales, por lo que la viabilidad de los cigotos será muy baja.
La translocación implica, como su nombre lo indica, un movimiento de un segmento de cromosoma a otro. Puede ser recíproca (o balanceada) cuando se intercambian segmentos con cromosomas no homólogos. Este tipo de mutación, al igual que la inversión no implica pérdida o ganancia de material genético, sino únicamente una reorganización y, normalmente, no tiene efectos sobre el fenotipo.
Cuando un fragmento de cromosoma se adhiere a otro, resulta una translocación no balanceada que, a menudo, trae consecuencias graves sobre el fenotipo. En el síndrome de Down familiar, uno de los progenitores tiene uno de los cromosomas 21 adherido al cromosoma 14; este individuo tiene un fenotipo normal aunque posee 45 cromosomas. Cuando ocurre la meiosis, una cuarta parte de sus gametos posee dos cromosomas 21, uno normal y el otro adherido al cromosoma 14. Cuando este gameto es fecundado, el resultado será un cigoto con 46 cromosomas, pero con tres copias del cromosoma 21: dos copias normales del cromosoma y una gran parte del otro adherido al cromosoma 14. Esto produce un desequilibrio genético que conduce a los cambios fenotípicos característicos del síndrome de Down común.
Las mutaciones genómicas se refieren a variaciones en el número de los cromosomas.
Esta variación se refiere a modificaciones que alteran a todo un juego de cromosomas o a la modificación en el número de un cromosoma particular.
Los organismos eucariontes presentan normalmente uno o varios juegos de cromosomas.
Normalmente, las especies eucariontes tienen dos juegos de cromosomas, por lo que se denominan diploides. Si un organismo presenta un juego adicional se denomina triploide o poliploide si posee más juegos. Los genetistas usan la letra n para representar un juego de cromosomas, por lo tanto, 2n significa diploide y 3n se refiere a un organismo triploide, etcétera.
La condición poliploide es muy común en plantas, pero es poco frecuente en animales; sin embargo, es relativamente común en lagartijas, anfibios y peces.
Los organismos normalmente diploides pueden tener células con diferente grado de poliploidía; por ejemplo, en las células hepáticas humanas pueden encontrarse núcleos triploides, tetraploides o aun octaploides, cuya función todavía no ha podido aclararse. Según la hipótesis más aceptada se trata de células cuyas necesidades de una determinada proteína sólo pueden satisfacerse con un mayor número de genes.
El ejemplo clásico de poliploidía se presenta en las glándulas salivales de la mosca de la fruta Drosophila melanogaster, que presenta varios ciclos de duplicación de ADN, sin división de citoplasma, dando lugar a los cromosomas politénicos, formados por las múltiples hebras de ADN dispuestas paralelamente. Este tipo de cromosomas ha sido de gran importancia para el estudio de la organización y funcionamiento de los cromosomas interfásicos.
En las plantas, el fenómeno de poliploidía es muy frecuente, encontrándose aproximadamente en el 30% de los helechos y de las fanerógamas. En la agricultura, su importancia ha sido enorme, pues casi todos los granos y frutos que se consumen son más grandes y resistentes, debido a su condición de poliploidía.
Igualmente, los floricultores toman ventajas de este fenómeno, pues las flores obtenidas son más grandes que sus originales diploides. Sin embargo, las plantas poliploides que poseen un número impar de juegos de cromosomas son usualmente estériles, pues no pueden producir gametos normales debido a la presencia de un número desigual de cromosomas. Por ejemplo, ciertos tipos de manzanas, algunas especies de patatas, las bananas cultivadas, los melones sin semillas y otras plantas más, son triploides y se propagan asexualmente.
El segundo tipo de mutación genómica, la aneuploidía, se refiere a la variación en el número de un cromosoma particular, es decir se gana o se pierde un solo cromosoma. Un organismo diploide que presenta aneuploidía puede poseer tres copias de un cromosoma particular en lugar de dos. En tal caso se dice que el individuo tiene una trisomía o que es trisómico. Si el organismo presenta un cromosoma de menos se dice que es monosómico.
La variación en el número de cromosomas resulta de un error que ocurre cuando los cromosomas homólogos que deben separarse durante la anafase de la primera división meiótica, no lo hacen o cuando las cromátidas hermanas no se separan durante la meiosis II o durante la mitosis, provocando con ello el fenómeno de no disyunción, que afecta la distribución correcta de los cromosomas en los gametos o la formación de mosaicos si el fenómeno ocurre en la mitosis. La aneuploidía causa, a menudo, la aparición de defectos fenotípicos severos, debido a que, en el transcurso de la evolución, se ha desarrollado una extraordinaria coordinación entre el genoma diploide, que hace que el individuo sólo pueda funcionar correctamente si están presentes las dos copias de los cromosomas, con excepción de la monosomía del cromosoma X que, a pesar de ser causante del aborto de muchos fetos, permite la supervivencia de otros individuos afectados.
La respuesta a este dilema se encuentra en la correlación entre la expresión de los genes y el número de cromosomas, pues el nivel de expresión de muchos genes está correlacionado con el número de genes por célula. Así, si una célula tiene tres copias de un cromosoma en lugar de dos, puede sintetizar 150% de la cantidad normal del producto de un gen; de la misma manera, si sólo tiene una copia sintetizará el 50%, en lugar del 100%, que se elaboraría con las dos copias normales. El desequilibrio en la expresión génica que se presenta en los individuos aneuploides resulta evidente, y explica por qué los portadores de estas mutaciones tienen menos oportunidades de sobrevivir. Sin embargo, es notable el hecho de que en las plantas los casos de aneuploidía, al igual que los casos de multiplicación de todo un juego de cromosomas, son más tolerados que en los animales.
Las alteraciones en el número de cromosomas en la especie humana ocurren frecuentemente durante la formación de gametos. Del 5 al 10% de las fecundaciones producen embriones defectuosos como resultado de anormalidades en el número de cromosomas y aproximadamente el 50% de los abortos espontáneos se deben a la presencia de anormalidades debidas a esta causa. Las aneuploidías más comunes son las trisomías de los cromosomas 21, 18 y 13 y las de los cromosomas sexuales. Se han encontrado trisomías que afectan a casi todos los cromosomas, con excepción del cromosoma 1, cuya trisomía posiblemente sea letal en estadios tan tempranos de desarrollo, que impida la implantación del embrión.
La edad de los padres parece jugar un papel importante en la aparición de anomalías en el número de cromosomas. Entre mayores sean los padres tienen mayor probabilidad de tener problemas de este tipo. Como en el síndrome de Down o trisomía 21, estudiado por Langdon Down en 1866, y cuya incidencia aumenta notablemente conforme avanza la edad de la madre. Ésta es la única trisomía autonómica que permite la supervivencia de los afectados mucho más allá del nacimiento, con la desventaja adicional de que la muerte de los adultos aquejados por el síndrome se debe a la enfermedad de Alzheimer, que los ataca a una edad más temprana que al resto de la población.